Evolutionary Biologist @ American Museum of Natural History
Research
I am a trans-disciplinary evolutionary biologist, and my main research interests are in the major transitions in evolution. These are the most significant events in the evolutionary history of life on Earth, the times at which the greatest biological innovations occurred.
The major transitions start at the emergence of life itself, and they include the origin of free-living cells, the evolution of complex cells (eukaryotes), the multiple origins of cooperation (from molecules at the origin of life all the way to insect and primate societies), and many more. A key focus of my research has been in investigating the role that membranes and membrane-bound proteins have played and continue to play in some of these transitions.
Membranes and their embedded proteins are crucial to most of life’s key processes, including respiration, cell-to-cell communication, reproduction, photosynthesis, vision, motion, feeding, energetics, and more. Membrane proteins amount to approximately one third of all proteins in the cell, and they compose about half of the membranes themselves. They are also the target of over half of all known drugs, so understanding their roles in the major transitions has implications beyond evolutionary biology.
I use mostly computational methods (bioinformatics and modelling) and theoretical analysis in my research, but I also do some work in the lab. I give a few details about my research below.
Adaptation to new environments
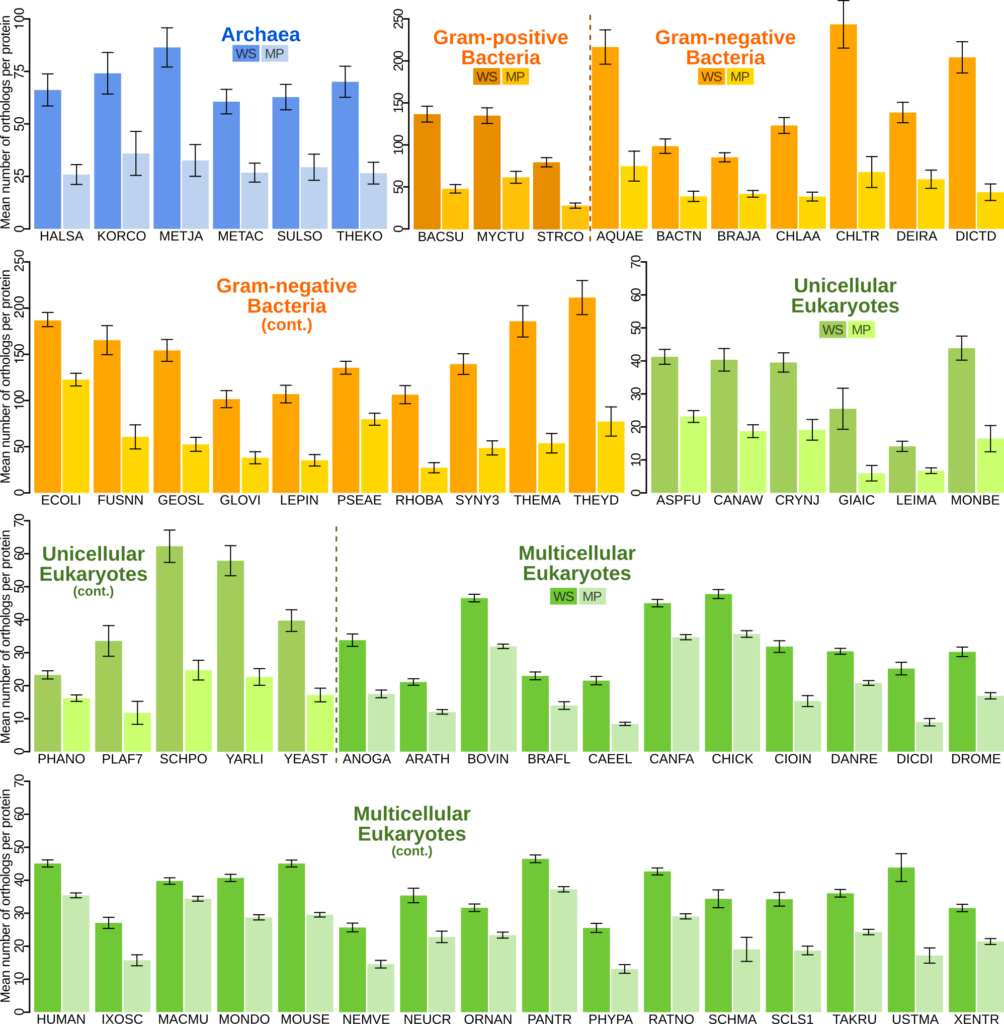
I detected that membrane-bound proteins are drastically less shared (they have fewer orthologs) than cytosolic proteins, across the tree of life. That is: you are far more likely to share a cytosolic protein than a membrane-bound protein with another species.
Membrane proteins are less shared. Across the tree of life, for every species I have studied, membrane proteins (MP) have fewer orthologues (they are shared with fewer species) than cytosolic water-soluble (WS) proteins.
Other authors had previously detected that membrane proteins evolve faster than cytosolic proteins, and that proteins exported outside the cell evolve fastest of all. Perhaps even more interestingly, the outside-facing portions of membrane proteins evolve faster than their inside-facing portions. However, it wasn’t known why.
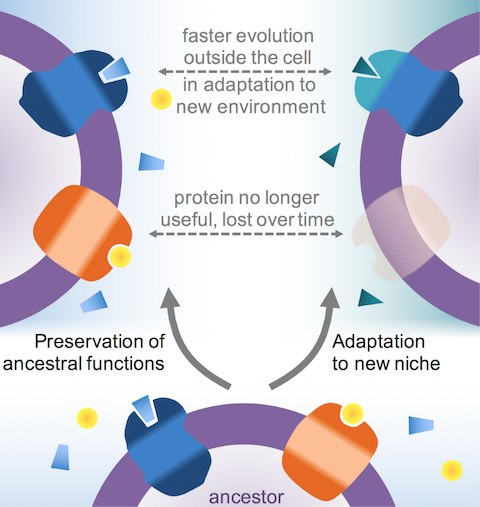
I suggested that the reason is adaptation to new niches. As cells come to colonise a new environment or specialise in a new function, it is the outside that experiences the strongest selective pressures. In this process, the outside of membrane proteins – as well as proteins exported from the cell – adapt and evolve faster than proteins and portions on the inside.
This explains why membrane proteins are less shared: in part, it’s because they evolve so much faster that when we compare them using the same criteria that we use to compare cytosolic proteins, we miss them. In some cases, we conclude that two proteins are not related because they don’t look similar enough, when the truth is that they are, they just evolve faster than what we’re used to. But I also showed that membrane proteins have indeed been lost. I explained that the reason is that some proteins are simply not useful when a cell gets a new function or adapts to a new environment .
Why membrane proteins may be shared less. In the colonisation of a new niche, some membrane proteins evolve beyond recognition (top), whereas some become useless and so are lost entirely (middle).
If I am right, then the effect should be strongest in prokaryotes, which have membrane proteins exposed to the environment, then a little weaker in unicellular eukaryotes, many of whose membranes are located in the membranes of organelles inside the cell, and weakest for multicellular eukaryotes, which have tissues and organs that provide additional layers of stability (homeostasis) for the cells to evolve in. Indeed, I have shown that this is the case.
You can read the paper discussing these findings here.
The origin of cells
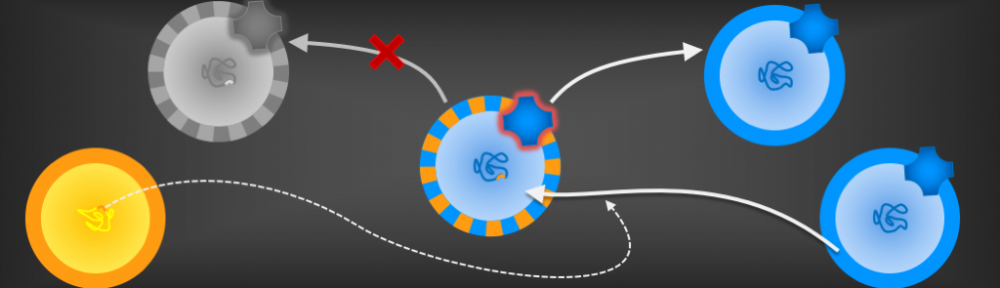
There are three types of organisms living on the Earth today: archaea, bacteria, and eukaryotes. We now know that archaea and bacteria (together the prokaryotes) are considerably older than eukaryotes, and they seem to be just as old as each other. Therefore, they are thought to have diverged from an organism fondly known as LUCA: the last universal common ancestor.
This organism was almost certainly cellular, just like archaea and bacteria are. We know this, because the two prokaryotic lineages share several crucial membrane-bound proteins, including the ATP synthase. So LUCA must have had a membrane.
But this presents a problem, because the membranes of archaea and bacteria are utterly different. So what were the membranes of LUCA like?
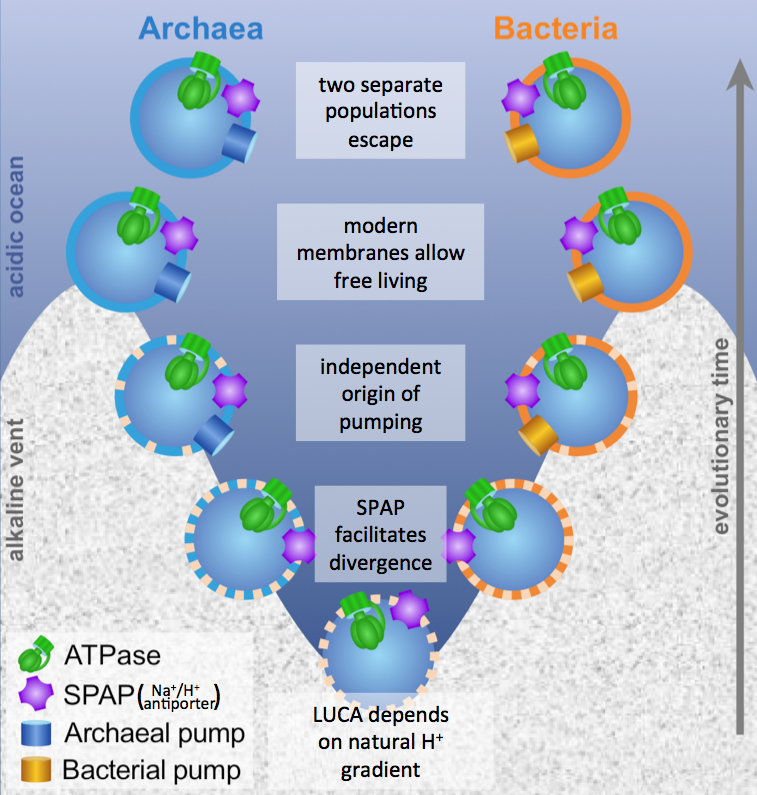
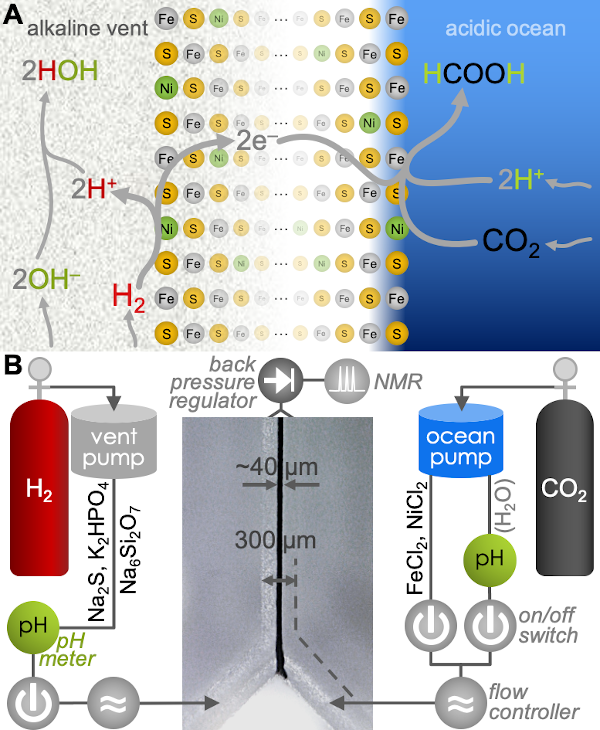
Using a mathematical model, I showed that the membranes had to be very leaky, unlike any membranes that modern cells have. This is because the ATP synthase, which archaea and bacteria share, needs to exploit an ion gradient (a high H+ or Na+ concentration outside, low concentration inside). However, archaea and bacteria don’t share the pumps that make those gradients (at least not ancestrally), so something else must have been making those gradients. Building upon previous work by – among others – Mike Russell, Bill Martin, and my own PhD supervisor Nick Lane, we suggested that LUCA’s cells were embedded right in the middle of a geologically generated H+ gradient in an alkaline hydrothermal vent (such as those in Lost City in the Atlantic Ocean). This allowed it to get the H+ from the acidic ocean side through an ancestor of the ATP synthase, but then neutralise them with OH– coming from the alkaline vent side.
A rock-bound LUCA. We built a model for the last universal common ancestor evolving in alkaline hydrothermal vents, accounting for the deep divergence in membranes and pumps between archaea and bacteria, but their similarity in the ATP synthase (ATPase) and sodium-proton antiporter (SPAP).
If this sounds a little strange to you, it does to me too. However, we need to explain why the membranes and pumps of archaea and bacteria are so different while their ATP synthases and (sodium-proton antiporter) are so similar. No other explanation I know of can account for this (which is why we set out to do this work in the first place).
With Nick’s group, we also wrote a review on the current standing of the alkaline vent theory for the origin of life. You can download it here.
Homochirality
The origin of life remains the most fundamental unresolved question in biology: the mystery of its own origin.
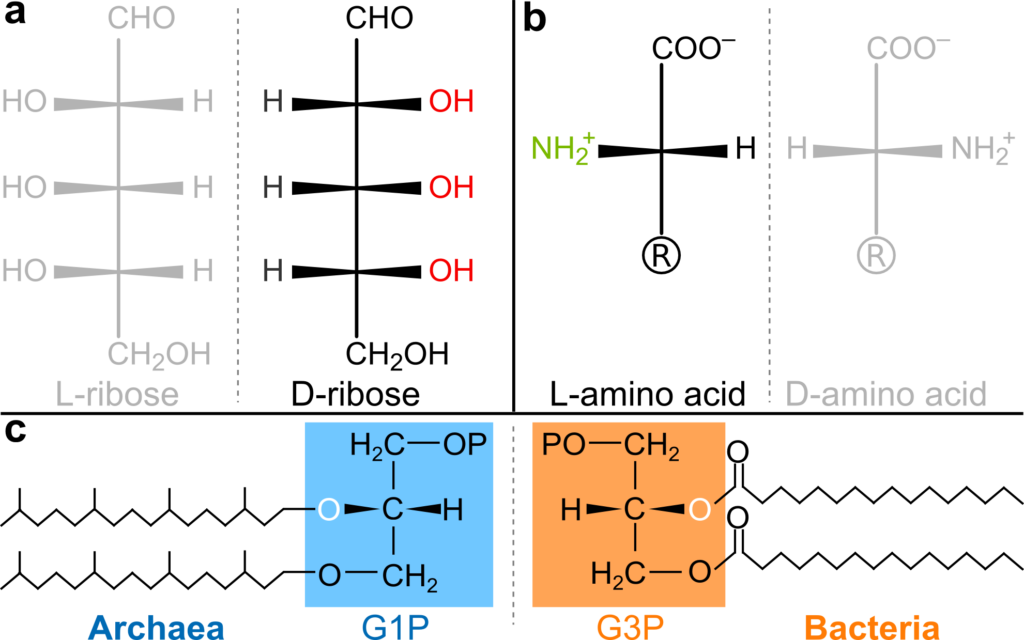
Amongst the many unsolved sub-questions is something called homochirality. There are two ways of making molecules such as the amino acids in proteins or the sugars in DNA. All known life on Earth chose only one of the two ways, and it is not clear how or why.
Homochirality in nature. Sugars in DNA (a) and amino acids in proteins (b) have only one of two possible orientations. However, there is a third case of dual homochirality in the phospholipid headgroups of phospholipids (c), which are fully homochiral in both archaea and bacteria, but with the opposite geometry.
Proposed causes range all the way from selective catalysis to polarised radiation due to imparity in the weak nuclear force in decaying stars. Since all of biology is already homochiral, the causes have understandably been sought mostly within physics or chemistry: what kind of external force could make one orientation decay faster, or the other one increase faster than their mirror images.
Enzymatic catalysis. Enzymes work in highly specific interactions with their substrates.
I have suggested, instead, that the reason stems from biology itself, and specifically from the fundamental difference between simple chemical and biological (enzymatic) catalysis.
Enzymes are highly specific, so a mirrored molecule, even though it may look very similar to a chemist’s eyes, is completely different for a three-dimensional enzyme that wraps around its substrate specifically.
As life arose and the first biological macromolecules started catalysing reactions, they had to choose one orientation. Catalysing a reaction for one chiral molecule is completely different from doing the same for its mirror image, so it’s only logical that life is homochiral. The way life does its chemistry would intrinsically lead to only one orientation.
You can read the full article here. I am presently designing a mathematical model to test some of these predictions.
The origin of life's first molecules
In this experimental work, funded by NASA and done in collaboration with Reuben Hudson at the College of the Atlantic in Maine (amongst several others), we addressed the the possible origin of carbon fixation at the emergence of life.
All living beings are built of organic molecules, which originate primarily from the reduction of CO2 through six known carbon-fixation pathways. Only one of these—the Wood-Ljungdahl acetyl-CoA pathway—is energetically profitable overall. Variations of it also happen to be present in both Archaea and Bacteria (thought to be the ancestral domains of the Tree of Life), which makes it immediately relevant to studies of the origin of life.
We used microfluidics to simulate geologically relevant, life-like microfluidic pH gradients across freshly deposited Fe(Ni)S (iron-nickel-sulphur) precipitates to demonstrate the first step of the Wood-Ljungdahl pathway: the otherwise unfavourable production of formate (HCOO–) from CO2 and H2. By separating CO2 and H2 into acidic and alkaline conditions—as they would have been in early-Earth alkaline hydrothermal vents—we demonstrated a mild, indirect, electrochemical mechanism of pH-driven carbon fixation. These results provide a potential bridge between geochemistry and biochemistry in the origin of carbon fixation at the emergence of life.
Beyond its immediate relevance to life’s emergence, the work has implications for both industrial and environmental chemistry.
Essentially every living being that we can see with the naked eye, from plants, to mushrooms, to sea kelp, to us animals, is a eukaryote. There are also dozens of unicellular lineages, but all eukaryotes have a host of very complex traits in common that are simply not present in prokaryotes (the archaea and bacteria).
Interestingly, the membranes of most archaea and bacteria are very different, but the membrane of eukaryotes look like those of bacteria. It would seem that eukaryotes, for some reason, changed their outer membrane from the archaeal type to the bacterial one, but nobody knows why.
There is multiple evidence that convincingly suggests that eukaryotes arose from a merger of bacterial cells into an archaeon. The internal endosymbiotic bacteria in time became what now are mitochondria, providing power for the whole cell in the form of ATP.
My present research aims to elucidate the reasons and circumstances of this process.